La historia evolutiva de los humanos modernos es una narrativa compleja y fascinante que ha sido objeto de estudio durante décadas. Gracias a los avances en tecnología genética y análisis computacionales, hemos logrado entender cada vez mejor cómo las poblaciones ancestrales se dividieron, migraron y se mezclaron a lo largo del tiempo. Un avance reciente en este campo ha sido el desarrollo de un modelo coalescente estructurado llamado cobraa, diseñado para detectar señales ancestrales profundas y complejas que no pueden ser captadas por modelos tradicionales. Este modelo ha revelado una estructura ancestral profunda compartida por todos los humanos modernos, marcada por la divergencia y posterior admixture de dos poblaciones ancestrales hace aproximadamente 1.5 millones de años.
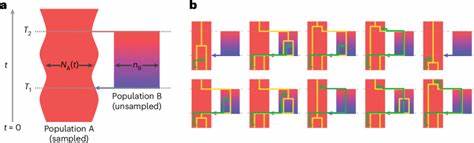
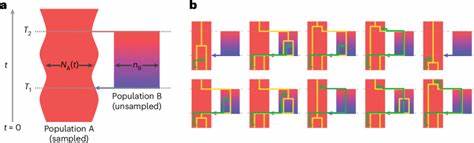
La coalescencia es un concepto fundamental en genética de poblaciones que permite rastrear hacia atrás en el tiempo el linaje común más reciente de alelos presentes en una población. Tradicionalmente, los modelos coalescentes han asumido que las poblaciones ancestrales se comportaban como panmícticas, es decir, con apareamiento aleatorio sin subestructuras internas. Sin embargo, este supuesto ha sido cuestionado debido a evidencias crecientes de que las poblaciones ancestrales humanas estuvieron estructuradas geográfica y genéticamente, con periodos de aislamiento y remezcla. Esto puede generar señales complejas en nuestros genomas que requieren modelos más sofisticados para su interpretación. El modelo cobraa incorpora explícitamente la posibilidad de una estructura poblacional ancestral, modelando dos poblaciones ancestrales (denominadas A y B) que divergieron hace alrededor de 1.
5 millones de años y que posteriormente experimentaron un evento de remezcla o admixture hace aproximadamente 300 mil años. Este acercamiento es significativo porque permite ir más allá de las asunciones simplificadas del panmixia al aprovechar la información contenida en las distribuciones condicionales de tiempos de coalescencia vecinos a lo largo del genoma, lo cual resulta en una mayor capacidad para identificar episodios históricos complejos de divergencia y mezcla. Los resultados de aplicar cobraa a datos genómicos de proyectos variados como el 1000 Genomes Project y el Human Genome Diversity Project son reveladores. Se detecta que todos los humanos modernos portan material genético derivado de estas dos poblaciones ancestrales en una proporción aproximada de 80% para la población mayoritaria A y 20% para la minoritaria B. Es importante destacar que tras su divergencia, la población mayoritaria A experimentó un fuerte cuello de botella poblacional, un fenómeno común en la historia evolutiva que implica una reducción drástica en el tamaño efectivo de una población, aumentando el efecto de la deriva genética y reduciendo diversidad.
Posteriormente, ambas poblaciones se combinaron mediante un evento de admixture, reconstituyendo la ascendencia común de los humanos actuales. El estudio ha sido capaz, gracias al modelo, de inferir cuáles regiones del genoma actual derivan de cada una de estas ancestrías antiguas. Se observa que el material genético proveniente de la población minoritaria B se encuentra con mayor frecuencia distante de regiones codificantes, lo que sugiere que pudo haber sido deletéreo en el contexto genético dominante de la población A y por tanto sujeto a selección negativa. Esta evidencia resalta que la interacción genética post-admixture no fue completamente neutral, sino que involucró dinámicas selectivas que moldearon la estructura genética que hoy observamos. Además, se establece una correlación significativa entre las regiones del genoma con ascendencia mayoritaria A y la divergencia con los genomas neandertales y denisovanos, dos ramas arcaicas del género Homo conocidas por su influencia en la genética de los humanos no africanos gracias a procesos de intercruce en el pasado.
Esto indica que la población A es probablemente la población ancestral común no solo de los humanos modernos, sino también de estos homínidos arcaicos, revelando conexiones profundas en la historia evolutiva de nuestro linaje. Este nuevo paradigma genera una visión más rica y dinámica de la evolución humana. La separación profunda y la posterior remezcla de estas ancestrías no solo ofrecen un contexto para entender los orígenes de Homo sapiens, sino que también explican patrones de diversidad genética que modelos anteriores no lograban capturar de manera satisfactoria. Cabe destacar que el porcentaje de admixture detectado (alrededor de 20%) es considerablemente mayor que el estimado para la contribución neandertal o denisovana en humanos modernos, y se encuentra distribuido en todas las poblaciones humanas actuales, por lo que no puede detectarse mediante análisis convencionales de estadística f. Por otro lado, el modelo cobraa no solo ha sido aplicado con éxito en humanos, sino que también se ha explorado su uso en otros mamíferos, como el delfín común y ciertas especies de primates.
En estos grupos, se identifican eventos estructurales y demográficos diversos, destacando la versatilidad y potencial del modelo para reconstruir historiales evolutivos complejos en distintas especies. En algunos casos, como en el murciélago particolorado, no se detectaron señales claras de estructura ancestral profunda, lo que puede servir para validar la sensibilidad del método y subrayar diferencias biológicas específicas entre especies. El modelo frente a los métodos tradicionales como PSMC (Pairwise Sequentially Markovian Coalescent) tiene ventajas sustantivas. Mientras que PSMC asume una población homogénea y puede producir inferencias erróneas o confusas cuando existen estructuras ancestrales, cobraa explota toda la información disponible sobre las transiciones entre tiempos de coalescencia adyacentes para distinguir entre cambios en tamaños efectivos poblacionales y la presencia de poblaciones distintas que existieron en el pasado. Esto aborda el llamado problema de no identifiabilidad que han enfrentado los modelos más simples.
De forma complementaria, al analizar las regiones genómicas asociadas con la entrada genética proveniente de la población minoritaria B mediante el uso avanzado de funciones de mapeo y anotación genómica, se identificaron categorías funcionales específicas que están enriquecidas. Las genes vinculadas a la adhesión celular neuronal, el reconocimiento neuronal y la transmisión de neurotransmisores resultan particularmente relevantes entre aquellas que muestran mayor presencia de material ancestro B. Esto sugiere que algunos elementos introgressados podrían haber influido en aspectos claves del desarrollo y la función cerebral, aportando potenciales ventajas o variantes funcionales a la especie. Al mismo tiempo, se detectaron categorías de genes donde la ascendencia B está significativamente escasa, vinculadas a procesos como la respuesta inmune adaptativa o la percepción sensorial, indicando que estas regiones podrían haber sido menos tolerantes a la introgression o que estuvieron sujetas a presión selectiva negativa, posiblemente debido a incompatibilidades genéticas o detrimento funcional. La contribución de este modelo y sus hallazgos va más allá de la reconstrucción puramente histórica.
Abre interrogantes importantes para la paleoantropología y la comprensión del contexto biológico de nuestros ancestros, proporcionando hipótesis fundamentadas que pueden relacionarse con evidencias fósiles y arqueológicas. En particular, la identificación de un período de estructura profunda coincidente con los tiempos estimados para la aparición de Homo sapiens anatómicamente modernos refuerza la conexión entre los cambios genéticos y los hechos paleoantropológicos. Además, el modelo ofrece herramientas valiosas para distinguir episodios de admixture estructurados de simples fluctuaciones en el tamaño poblacional, una distinción crucial para interpretar correctamente eventos complejos en la evolución humana y evitar conclusiones erróneas sobre migraciones o contactos entre grupos. En conclusión, el desarrollo y aplicación del modelo coalescente estructurado cobraa marca un avance significativo en la comprensión de la evolución humana. Al revelar la existencia de una estructura ancestral profunda compartida por todos los humanos modernos, desvela la historia dinámica y mezclada de nuestros antepasados, y cómo estos eventos moldearon los genomas que hoy portamos.
A medida que la tecnología genómica y los modelos computacionales continúen evolucionando, este tipo de enfoques promete seguir ampliando nuestro entendimiento sobre la historia hereditaria y adaptativa del ser humano y otras especies.